|
Environmental Chemistry Letters |
|
© Springer-Verlag 2007 |
|
10.1007/s10311-007-0100-7 |
Original Paper
Recycling greenhouse gas fossil fuel emissions
into low radiocarbon food products to reduce human genetic damage
Christopher P. Williams1 ![]()
|
Radiocarb Genetics, |
|
|
Christopher P. Williams |
Received: 12 February 2007 Accepted:
15 March 2007 Published online: 23 May 2007
Abstract Radiocarbon from nuclear fallout is a known health risk. However, corresponding risks from natural background radiocarbon incorporated directly into human genetic material have not been fully appreciated. Here we show that the average person will experience between 3.4 × 1010 and 3.4 × 1011 lifetime chromosomal damage events from natural background radiocarbon incorporated into DNA and histones, potentially leading to cancer, birth defects, or accelerated aging. This human genetic damage can be significantly reduced using low radiocarbon foods produced by growing plants in CO2 recycled from ordinary industrial greenhouse gas fossil fuel emissions, providing additional incentive for the carbon sequestration.
Keywords Radiocarbon - Mutations - DNA - Histones - Carbon dioxide - Sequestration
Introduction
At first glance, radiocarbon-induced genetic damage in humans and global climate change from fossil fuel greenhouse gas emissions appear to be entirely unrelated problems. But these disparate issues share a surprising environmental interconnection, which until now has been overlooked, and which offers the possibility of addressing them both with a single, novel approach–recycle a seemingly useless and environmentally harmful industrial waste material into heretofore unavailable low radiocarbon food products.
Natural background radiocarbon
Libby and his coworkers were the first to recognize the significance of radiocarbon in the environment (Anderson 1947; Libby 1960). Approximately, one in 750 billion carbon atoms in the biosphere (that is, the atmosphere, oceans, and living organisms) is radioactive carbon-14, or radiocarbon, with a half-life of 5,730 years. The natural background level of radiocarbon is in general the result of new 14C produced from cosmic ray interaction with 14N in the upper atmosphere achieving equilibrium with the radioactive decay of 14C back into 14N. Libby found that even this trace amount of natural background radiocarbon resulted in a decay rate of 15.367 dpm g−1 for ordinary carbon. Because photosynthetic plants and microorganisms use atmospheric and aqueous CO2 containing this radiocarbon, all living organisms that derive their food directly or indirectly from plants, including man, contain this same natural background level of radiocarbon. Libby realized that once an organism died and failed to take in fresh carbon containing the natural background level of radiocarbon, its radiocarbon content would steadily drop by half every 5,730 years. His Nobel Prize-winning radiocarbon dating method has proven invaluable in archeology and anthropology to estimate ages of organic materials less than 100,000 years old. Organic material more than 100,000 years old, including coal, oil, and natural gas deposits, will have lost more than 99.999% of any original radiocarbon and for practical purposes can be considered radiocarbon free.
But radiocarbon decay does not only occur in dead organisms. Since carbon makes up about 18% of the human body by weight, an average 150 lb, adult contains about 6.2 × 1026 carbon atoms, of which 1 in 750 billion, or 8.2 × 1014, are carbon-14. Such an individual therefore experiences about 3,150 radiocarbon decay events every second, 99 billion per year, and 6.8 trillion over an average 77.9 year lifetime (adjusting for fewer cells during the growing years), due solely to natural background radiocarbon.
More than 40 years ago, near the height of atmospheric nuclear weapons testing, Linus Pauling sounded an alarm concerning the health risks from radiocarbon in fallout, which nearly doubled the natural background level of radiocarbon in the mid-1960s (Pauling 1958; Pauling 1963). He estimated that general carbon-14 beta decay from this fallout, would ultimately be responsible for one and a half million viable children with gross physical or mental birth defects, and for nearly 15 million embryonic, neonatal, and childhood deaths. Within a few years after the signing of the “Treaty banning nuclear weapon tests in the atmosphere, in outer space and under water” in 1963, atmospheric radiocarbon began to fall back to its previous level, as did public concern over natural background radiocarbon.
But if doubling the natural background level of radiocarbon would cause such huge human health risks, why wasn’t the natural background baseline itself considered hazardous? Perhaps the baseline natural background radiocarbon level was simply accepted as an unavoidable natural hazard. There may also have been belief in an implicit (and unproven) assumption that the detrimental effects of any form of radiation at or below certain levels were non-linear, and in particular sub-linear (i.e., that below a certain lower threshold it was completely harmless, or at least much less harmful), and that the natural background level of radiocarbon fell within that safe region. But a National Academy of Sciences panel recently completed a 5 year study which concluded that the evidence best supports a “linear no threshold” (LNT) model for extremely low doses of radiation (Board on radiation effects research 2006). The LNT model states there is no safe lower level or threshold for radiation, and that any amount, no matter how small, is potentially damaging to health. The panel also made particular note that radiation can alter the molecular structure of DNA, and that “some of these molecular changes are so complex that it may be difficult for the body’s repair mechanisms to mend them correctly.”
Radiocarbon in food is not unavoidable. One of the major environmental and political issues of our day is how to avert or minimize the potentially devastating effects of global warming. A leading option to reduce atmospheric carbon dioxide, the greenhouse gas believed most responsible for the anthropogenic heating of the earth’s atmosphere, is to capture or sequester it from industrial fossil fuel combustion gas emissions. But carbon sequestration processes are quite expensive, and could produce billions of tons of useless and potentially harmful waste material each year––waste that would need to be permanently dumped deep into the oceans or abandoned mines and aquifers. Not surprisingly such efforts face considerable opposition. Finding a significant new use for carbon dioxide, and particularly low radiocarbon CO2 extracted from fossil fuel emissions or obtained as a byproduct from coal gasification or the refining of oil, could profoundly advance carbon sequestration efforts. One option deserving consideration is using this material for growing safer, low radiocarbon food.
Theory
and calculations
Pauling based his
estimates for genetic damage from radiocarbon solely on general ionizing
radiation arising from beta decay of all the carbon-14 scattered throughout the
human body. However, in those particular cases where a radiocarbon atom
incorporated into the genes undergoes decay, two additional mechanisms can
produce genetic damage. The loss of a 156 kev
beta particle from the nucleus during radiocarbon decay causes the
transmutation of that carbon-14 atom to the chemically different nitrogen-14.
The beta emission is also coupled with powerful recoil of the residual nitrogen-14,
which will in most cases completely rip the atom out of its original molecule
(the “Szilard-Chalmers Effect”). Asimov (1955)
was the first to recognize that this transmutation and recoil effect guarantee
that any carbon-14 decay occurring within the 39 different carbon atoms in the
four DNA nucleotides (see Fig. 1)
will produce a chemical change, or mutation. Pauling was also aware of this
“special mutagenic action of carbon-14.” However, based on the limited
knowledge of genetics available at the time (the genetic code had not even been
worked out), he calculated that this special mechanism would amount to less
than 10% of the genetic damage caused by general ionizing radiation from
carbon-14, and was therefore not considered particularly significant.
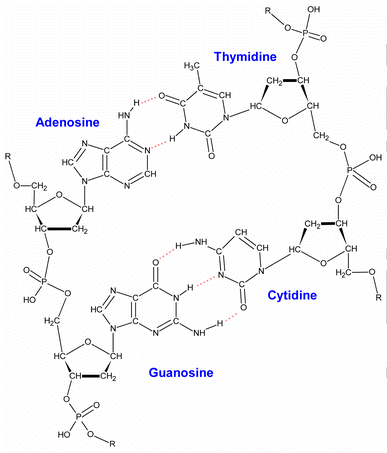
Fig. 1 Possible DNA carbon positions. Radiocarbon atoms
incorporated at any of these different nucleotide carbon positions may cause
irreparable damage when they decay
Pauling’s (1958)
estimates assumed there were 50,000 genes in humans and 200,000 carbon atoms
per gene, for a total of 1.0 × 1010 carbon atoms per cell.
Although for its time this was an excellent estimate for the protein coding
regions of the genome, it overlooked the rest of the genome, which contains
regions now known to be critical to genetic regulation or the structural
integrity of the chromosome.
We now know that human cells (with the
exception of non-nucleated cells such as red blood cells) each have about
6.4 × 109 nucleotide base pairs (diploid number), or a
total of 1.28 × 1010 nucleotide bases per cell. Of the
four DNA nucleotides, three (adenosine, guanosine, and thymidine) have ten
carbons each and the other (cytidine) has nine. Since the overall human AT and
GC content is about 59 and 41%, respectively, this gives an average of 9.8
carbons per human nucleotide. Thus the chromosomal DNA in every nucleated human
cell contains about 1.25 × 1011 carbon atoms, more than 12
times Pauling’s estimate. Approximately one in six human cells contains a
radiocarbon atom incorporated somewhere in its chromosomal DNA.
In addition to DNA in the chromosomes, human
cells contain extra-chromosomal DNA in the mitochondria, which are critical to
cellular respiration. Each mitochondrial DNA (mtDNA) molecule is 16,569 bp long, with a variable copy number, typically between
1,000 and 10,000, per human cell. The total amount of mtDNA is only about 1.3%
that of chromosomal DNA, but since mtDNA has less efficient repair mechanisms
than chromosomal DNA, its damage by radiocarbon will have a greater tendency to
go unrepaired. The high copy number of mtDNA allows more of such individual
mutations to be tolerated. But as they accumulate in germ cells over many
generations, such errors are more likely to afflict subsequent offspring.
Indeed, numerous inherited mitochondrial disorders are known to exist (though
their actual origins are unknown).
Furthermore, Pauling’s calculations of
chromosomal damage do not take into account the significance of carbon-14 in the
histone proteins. In addition to DNA, the chromatin material of chromosomes
also contains special proteins around which the DNA is tightly packed. These
histone proteins are strongly basic (due to excess lysine and arginine
residues) and thus neutralize the DNA’s acidity (due to its linking phosphate
groups). If the entire DNA in a single microscopic human cell were stretched
out linearly, it would be almost 2 m (6 feet) long. Histones allow
this large amount of DNA to be highly coiled and compacted in domains known as
nucleosomes so the entire complement of human DNA will fit within a 10 μm cell nucleus less than 1/500,000th of its fully
extended length. Incredibly, this highly coiled and compacted DNA retains full
functionality and can be accurately replicated and transcribed as needed.
Although the precise structure of chromatin varies according to tissue type,
the position within the chromosome, and point of time within the cell cycle, on
average every 200 bp of DNA is associated with a
histone nucleosome core particle consisting of two molecules each of histones
H2A, H2B, H3, and H4 along with a single histone H1 linker. Through extremely
complex enzymatically controlled processes involving activities such as
methylation and acetylation of specific basic histone residues, the cell is
able to unwind and open up the correct sections of DNA precisely when needed
for replication or for transcribing genes into messenger RNA. So not only are
histones required for ultra compact DNA storage, they also are integrally
involved in DNA processing. This critical role for histones most likely
explains why their amino acid sequences are among the most highly conserved of
all known proteins.
A carbon-14 decay that occurs within the
histone protein portion of chromatin may not necessarily cause a direct DNA
mutation. However, since the histone protein is in such close proximity to the
DNA, there is a significant chance that the beta particle emitted during
carbon-14 decay in a histone protein will directly strike the DNA.
Additionally, the chemically modified amino acid residue (after the
transmutation of carbon-14 to nitrogen-14) in the histone protein itself is
likely to be highly reactive, or to generate reactive free radicals species
nearby. Because of the close proximity to the DNA, the chances of these
reactive chemical species interacting with a DNA base and causing a mutation
are significant. But even without actually damaging the DNA directly, a
carbon-14 damaged histone protein itself may fold differently, or become
cross-linked to itself or to an adjacent protein, or have a key residue
modified. Any of these or other possible scenarios that cause structural
alteration of a histone protein could result in abnormal cellular behavior,
whether by preventing unfolding of the DNA when required, or by causing the
unfolding to occur when it should not, or via some other as yet undefined
mechanisms.
Because we know the amino acid sequences of
the typical histone proteins and the general structure of nucleosomes, we can calculate
that there are roughly 1.47 times as many carbon atoms in the histone proteins
of the chromatin as there are in the DNA itself. Approximately one in four
human cells contains a radiocarbon atom incorporated somewhere within its
histone proteins.
Estimates for the number of nucleated cells
in the average adult human body generally range from 1013 to 1014
cells. Using the most conservative estimate of 1013 cells, it can be
calculated that the human body contains at least 1.25 × 1024
total carbon atoms within the chromosomal DNA, of which 1.67 × 1012
are carbon-14 atoms, and 1.84 × 1024 total carbon atoms
within its histones, of which 2.46 × 1012 are carbon-14
atoms.
The number (N)
of atoms that will decay during any time period (t) is given by the standard
equation for radioactive decay:
|
|
where
|
|
N 0 = initial number of carbon-14 atoms |
|
|
λ = 0.693/t 1/2 |
|
|
t 1/2 = carbon-14 half-life (5,730 years, or 1.808 × 1011 s) |
|
|
t = time period |
Applying this equation
for radioactive decay to the amount of radiocarbon incorporated in the various
genetic materials gives us estimates of the cumulative number of human genetic
damage events for various time intervals (see Table 1).
Table 1 Cumulative human genetic damage events from natural
background radiocarbon
|
Time Interval |
Chromosomal DNA |
mtDNA |
Histones |
Total |
|
1 s |
6.4 |
0.083 |
9.4 |
16 |
|
1 min |
380 |
5.0 |
570 |
950 |
|
1 h |
23,000 |
300 |
34,000 |
57,000 |
|
1 day |
550,000 |
7,200 |
810,000 |
1,400,000 |
|
1 year |
200,000,000 |
2,600,000 |
300,000,000 |
500,000,000 |
|
Lifetime |
14,000,000,000 |
180,000,000 |
20,000,000,000 |
34,000,000,000 |
The
calculated number (within two significant digits) of radiocarbon decay events
within each genetic component is based on the low end estimate of
10 trillion total human cells. mtDNA values assume 5,000 mtDNA copies per
cell. Lifetime values are based on the 77.9 year average
Results
and discussion
Although Pauling and other scientists were justly concerned about severe health risks from increasing the natural background level of radiocarbon, overlooked until now were the harmful health effects arising from the baseline natural background radiocarbon itself, and the subsequent benefits that might result if such levels could be reduced.
Our calculations show that an average person whose body contains 1013 cells will experience at least 3.4 × 1010 lifetime chromosomal damage events solely from natural background radiocarbon incorporated into DNA and histones. These are likely to be low-end estimates. The actual numbers could be as much as ten times higher if the average human body contains as many as 1014 cells. This significant amount of cumulative lifetime genetic damage should not be quickly dismissed as inconsequential, especially if the currently accepted LNT model for radiation damage is true and there is no safe lower level or threshold for radiation.
The total number of carbon atoms in the cell closely associated with genetic activity, including chromosomal DNA, histones and mitochondrial DNA, is more than 30 times greater than that originally estimated by Pauling. Thus, rather than damage from the special effect of carbon-14 in the genetic material being only a negligible 10% of the overall damage arising from carbon-14 beta decay, it could be as much as three times greater than general carbon-14 beta decay. This would nearly quadruple Pauling’s already high (1963) estimates for overall human suffering resulting from atmospheric nuclear weapons testing. The number of grossly deformed children could go from 100,000 to 400,000, and the number of stillbirths and childhood deaths from 15 to 60 million.
Pauling also implicitly equated the severity of individual mutations arising from general ionizing radiation from beta decay of carbons outside the DNA to those caused by carbon-14 decay in the carbons within the DNA itself. This may not be true.
Human beings (as well as most other organisms) have complex systems for different types of DNA repair (Wood et al. 2001; Sancar et al. 2004), and not all mutations can be as successfully detected or repaired. For example, some mutations, such as thymine dimerization induced by overexposure to UV radiation, or the oxidation of a nucleic acid base by an oxidant, may involve no loss of DNA sequence information, and can be enzymatically repaired with near 100% efficiency. Likewise, other mutations that remove or modify a single DNA base, such as may occur from a free radical produced by beta decay, can also potentially be repaired with near 100% efficiency using the backup information from the complementary base on the opposite DNA strand.
However, there are three reasons why more severe mutations affecting multiple residues may be expected when a radiocarbon incorporated in a DNA nucleotide itself (see Fig. 1) undergoes decay as compared to a mutation from stray ionizing radiation. First, the high-energy beta particle is emitted from within the genetic material itself, and thus has a much higher probability of striking multiple nearby DNA residues or forming free radicals in their vicinity than does a beta particle originating outside the chromosome. Second, there occurs a transmutation of carbon-14 to nitrogen-14, ensuring a significant chemical change in the affected DNA residue. Third, the beta emission produces a severe recoil in the new nitrogen atom, which is likely to result in a nitrogen free radical and to further increase the chance that the remaining nucleotide residue is converted into some highly reactive species. Any of these reactive species produced could subsequently attack adjacent nucleotide bases. Such complex damage involving multiple residues is more likely to be either unrepairable or else prone to erroneous repair. Overall long term damage and health risks are more closely related not to the total number of mutations which initially occur, but rather to the number of those that are ultimately unrepairable. In addition to birth defects and cancer, unrepaired genetic damage may also lead to genetic diseases and accelerated aging (Sancar et al. 2004; Park and Gerson 2005).
Significant reductions in human radiocarbon levels are theoretically possible using low radiocarbon foods produced from ancient subterranean sources of carbon, such as fossil fuels, which are known to be almost completely free of radiocarbon. The first such proposal involved the growing of low radiocarbon food in greenhouses or other closed systems using carefully mined coal and special handling processes (Matthews 1995). A subsequent variation was the use of ordinary greenhouses, alternatively using thermally degraded limestone (calcium carbonate) or directly burning fossil fuels on site to provide the requisite low radiocarbon CO2 (Miekka and Mackie 1999). To date there does not appear to have been any actual commercial application of these methods. This may be partially due to failure to fully recognize the potential damaging effects of radiocarbon as calculated here, and partially due to the apparent costs and difficulty of these specialized methods for producing low radiocarbon CO2. However, a better alternative exists.
More than 27 billion (2.7 × 1010) metric tons, or nearly 4 tons for every person in the world, of low radiocarbon CO2 fossil fuel emissions from human activities are released into the atmosphere each year. These greenhouse gas emissions are generally considered the primary contributors to global warming and potential climate change (Oreskes 2004). Finding a safe and inexpensive way to minimize these CO2 greenhouse gas emissions is one of today’s most pressing political and environmental issues. Nearly all discussions involving CO2 sequestration consider this material relatively useless and simply a waste that requires permanent disposal. Large scale recycling of fossil fuel CO2 emissions as a means to grow safer, radiocarbon-reduced foods has not been considered until now and deserves further consideration. It is interesting to note that fossil fuel combustion gas emissions are already producing significantly radiocarbon-depleted vegetation, albeit unintentionally, with radiocarbon reductions of 12% or more found in grasses collected within a few meters of heavily trafficked French highways (Lichtfouse et al. 2005).
Further research involving mass spectrometry of small double stranded oligonucleotides or peptides enriched with radiocarbon could reveal the specific types of biochemical damage this substance can cause in DNA and histones. Studies using mice or other organisms with cohorts fed diets having normal (natural background), reduced, and elevated levels of radiocarbon could ascertain its overall impact on rates of cancer, birth defects, and aging. Although a low radiocarbon diet would be expected to primarily benefit organisms during their maximal periods of growth when new cells and DNA are being formed, studies are also needed to see if mature individuals could receive any benefit from a low radiocarbon diet.
Because it takes carbon materials nearly six half-lives, or more than 32,000 years, for their radiocarbon levels to drop by 98%, for all practical purposes we must consider fossil fuels and fossil fuel emissions a non-renewable low radiocarbon resource. Admittedly limestone and other ancient carbonate rocks are also potential sources of low radiocarbon CO2, but they would likely be considerably more expensive and would only exacerbate the problem of rising atmospheric CO2. If low radiocarbon food is found to be beneficial to human health, for the sake of future generations we should not waste the readily available low radiocarbon CO2 in fossil fuel emissions by irretrievably releasing it to the atmosphere.
It also should be noted that the total amount of natural background radiocarbon in the atmosphere and in all land organisms is only about 3,200 kg, or 7,000 lbs. (Pauling 1958). That is why a relatively small amount, just 74 kg, of radiocarbon produced from the atmospheric testing of just one 30 megaton nuclear weapon could increase the worldwide atmospheric radiocarbon level by 2.3%. Although not pleasant to consider, an additional benefit of stockpiling low radiocarbon CO2 in a recoverable manner is to provide for a safe emergency world food supply in the event of a catastrophe, such as a nuclear exchange or even a terrorist action, which could appreciably increase the background level of radiocarbon in the atmosphere for many years.
Conclusion
Low radiocarbon foods produced using CO2 sequestered from fossil fuel emissions provide a means to reduce human chromosomal damage from natural background radiocarbon as well as from potentially elevated levels of radiocarbon resulting from future atmospheric nuclear explosions. Should a low radiocarbon diet ultimately prove to offer significant human health benefits, this would not only boost the current incentive for carbon sequestration of fossil fuel emissions beyond that of simply reducing the greenhouse gas effect, but would also shift carbon sequestration technologies toward methods which permit the subsequent cost effective recovery and reuse of low radiocarbon CO2 for food production.
|
Anderson EC, et al (1947) Radiocarbon from cosmic radiation. Science
105:576–577 |
|
|
|
Asimov I (1955) The radioactivity of the human body. J Chem Educ 32:84–85 |
|
|
|
Board on radiation effects research (2006) Health risks from exposure
to low levels of ionizing radiation: BEIR VII phase 2. National Academies
Press, Washington |
|
|
|
Libby, WF Radiocarbon dating. Nobel Lecture (1960) (http://www.nobelprize.org/nobel_prizes/chemistry/laureates/1960/libby-lecture.pdf)
|
|
|
|
Lichtfouse E, Lichtfouse M, Kashgarian M, Bol R (2005) 14C
of grasses as an indicator of fossil fuel CO2 pollution. Environ Chem Lett 3:78–81 |
|
|
|
Matthews M (1995) Method for improving the chemical stability of living
organisms. US Patent No. 5,471,785 |
|
|
|
Miekka F, Mackie P (1999) Methods and apparatus for reducing
carbon 14 in living tissue. US Patent No. 5,956,896 |
|
|
|
Oreskes N (2004) The scientific consensus on climate change.
Science 306:1686 |
|
|
|
Park Y, Gerson SL (2005) DNA repair defects
in stem cell function and aging. Annu Rev Med
56:495–508 |
|
|
|
Pauling L (1958) Genetic and somatic effects of carbon-14. Science
128:1183–1186 |
|
|
|
Pauling L (1963) Science and peace. Nobel Lecture (http://nobelprize.org/nobel_prizes/peace/laureates/1962/pauling-lecture.html)
|
|
|
|
Sancar A, Lindsey-Boltz LA, Keziban U, Linn S (2004) Molecular mechanisms of
mammalian DNA repair and the DNA damage checkpoints. Annu
Rev Biochem 73:39–85 |
|
|
|
|
|
|